Гипотеза о механизмах памяти
Гипотеза о механизмах памяти
Трудно предложить гипотезу о механизмах памяти, пригодную для алгоритма интеллекта. Попробую изложить свою попытку. Для обозначения моделей и их сочетаний я буду пользоваться следующими заимствованными из лингвистики терминами. «Буква» — элементарная модель, самый малый значимый признак. «Алфавит» — совокупность буквмоделей, формируемых одним типом рецепторов; «алфавитов» может быть много: световые, звуковые и др. (см. ниже) «Слово» — более сложная модель, состоящая из элементарных моделей, то есть «букв», но очень хорошо организованная и выступающая как одно целое. «Фраза» — соединение «слов», чаще всего временное и непрочное. «Буква обобщенности» —знак, указывающий уровень обобщения моделей, их место в «иерархии блочности».
Модели и ансамбли нейронов. У человека в коре очень много нейронов, их количество оценивают в 10 и более миллиардов. Каждый нейрон соединяется с многими сотнями других. Таким образом, имеются почти беспредельные возможности для образования структур из нейронов, объединенных проходимыми («проторенными») связями. Такие структуры — нейронные ансамбли — могут выступать в качестве моделей объектов внешнего мира. Один нейрон может включаться в несколько ансамблей, как это показано на рис. 15. Модель повторно возбуждается («вспоминается»), если возбудится некоторый процент входящих в ее ансамбль нейронов. Почему не возбуждаются все нейроны коры. Ведь связей между ними вполне достаточно. Чтобы ответить на этот вопрос, нужно сделать еще одно предположение: когда один нейрон возбуждается, соседние тормозятся. Этот принцип «индуктивного торможения» установлен И. П. Павловым. Таким образом, если допустить, что 30% нейронов данной модели возбудятся извне, то все окружающие должны бы затормозиться. Так и происходит, но связи к остальным 70% нейронов данной модели настолько проторены, что эти нейроны больше возбуждаются по связям, чем тормозятся «по площади». В результате активируется весь ансамбль, вся модель, а другие модели остаются заторможенными, несмотря на то что часть возбужденных в данной модели нейронов входит и в них. Такова гипотеза об ансамблях нейронов.
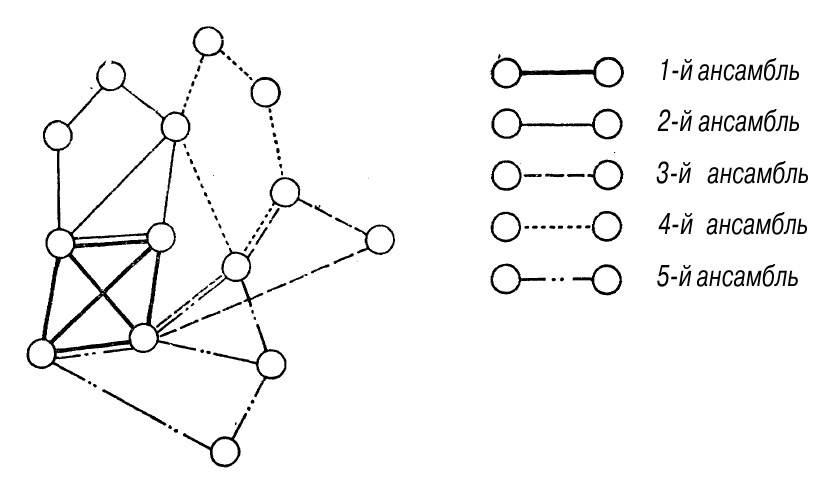
Рис. 15. Нейронные ансамбли.
Для проверки этой гипотезы сотрудниками отдела биокибернетики разработан макет нейронной сети, содержащий 22 узла и наборное поле, которое позволяет осуществлять все возможные соединения между узлами. Узлы (модели нейронов) представляют собой усилители постоянного тока с нелинейной (S-образной) характеристикой. В качестве связей использованы постоянные резисторы. Вес связи считается обратно пропорциональным сопротивлению резистора.
Макет создавался для получения ответа на следующие основные вопросы:
— можно ли технически реализовать устойчиво работающую сеть из нейронных ансамблей так, чтобы при возбуждении одного ансамбля не возбуждались одновременно и другие, связанные с ним; — можно ли построить устойчивую сеть, если ансамбли пересекаются, то есть одни и те же узлы входят в состав различных ансамблей; — можно ли добиться четкого перехода возбуждения с одного ансамбля на другой, если они пересекаются значительной частью своих узлов.
Эксперименты с макетом дали положительные ответы на все эти вопросы. Оказалось, однако, что для успешной работы необходимо использовать централизованную систему, управляющую активностью всей сети. По своим свойствам и назначению эта система аналогична системе усиления-торможения (СУТ), которая подробно будет описана ниже. Следует отметить, что многие исследователи, пытавшиеся построить устойчивую ансамблевую сеть без централизованной системы управления активностью, встретились с большими трудностями и вынуждены были накладывать жесткие ограничения на допустимую структуру сети.
На макете было также показано, что устойчиво работающая ансамблевая сеть может иметь число ансамблей, превышающее число узлов сети. Поэтому в некоторых задачах, используя ансамблевые сети, можно получить более экономную аппаратурную реализацию. С увеличением числа узлов эта экономия становится все более ощутимой. К сожалению, построение ансамблевых сетей большого объема связано пока со значительными трудностями.
Устройство «рецепторного поля». На рис. 16 показана гипотетическая схема «рецепторного поля», объясняющая принцип совмещения временной и постоянной памяти. Такое совмещение, несомненно, имеет место в коре мозга.
Система рецепторов Рц подает активность на «рецепторное поле», состоящее из довольно большого количества кадров, построенных так, что в каждом из них имеются представительства — элементы для каждого рецептора. Для удобства понимания кадры расположены в виде кольца. Число кадров значительное, но не бесконечное. Предположим, что есть переключатель в центре кольца, который или поворачивает его на один кадр в каждый такт времени, или поочередно переключает связи от рецепторов с одного кадра на другой, соседний. Имеется настройка кадров Я, обеспечивающая фокусирование, то есть позволяющая им четко воспринимать избранную деталь и неясно видеть весь объект. Она же передвигает фокус по структуре объекта. Еще одно условие: настройка «устает», поэтому после нескольких тактов рецептор отключается совсем.
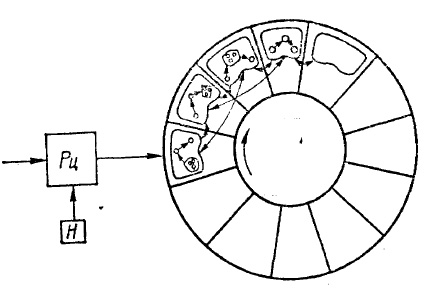
Рис. 16. Схема «рецепторного поля»: Рц — рецепторы, Н — настройка.
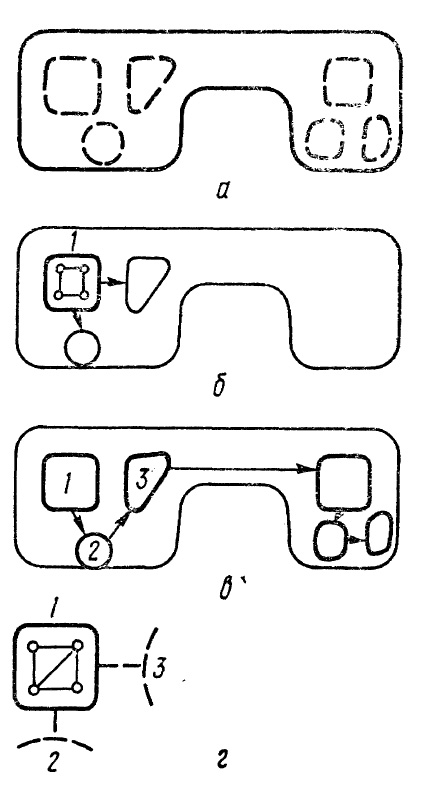
Рис. 17. Система моделей разной степени обобщенности, отражающая объект.
Характеристики затухания активности элементов очередного кадра после отключения от них рецептора имеют вид, показанный на рис. 8. Затухание активности в самом первом кадре происходит раньше, чем завершится полный круг переключения.
Картины, представленные в кадрах, показаны на рис. 17. В первый такт времени система рецепторов при отсутствии фокусировки неясно воспринимает объект как целое, и он отражается в виде наиболее обобщенной модели (рис. 17, а). Между активированными элементами возникают связи. Предположим, что объект «заинтересовал» интеллект (об этом — ниже), тогда в следующий такт, приходящийся на следующий кадр, зрением, умеренно сфокусированным на верхнем левом углу, преимущественно воспринимается часть объекта, обозначенная цифрой 1. Создается временная модель этой части, представляющая собой не очень четкое воспроизведение последней при обобщении всего остального объекта (рис. 17, б). Затем фокусировка переходит на часть 2, далее на часть 3 объекта, и они отпечатываются в модели. Модели в предыдущих кадрах еще сохраняют активность, и от них проторяются связи к следующему кадру. В каждом кадре на соответствующих элементах отмечаются направление и степень фокусировки всей системы рецепторов (назовем ее «глазом»). После того как настройка на крупные части обойдет их и они отпечатаются на кадрах, структура их расположения вырисуется в «модели-схеме» (рис. 17, в). Вся серия картин, отображенных в кадрах, объединена продольными связями. На этом восприятие может закончиться, но может быть продолжено на следующий цикл, состоящий в еще большей фокусировке (напряжении) «глаза», нацеленной на детальное рассматривание каждого блока с отражением тонких подробностей его структуры, «привязанных» к блоку, а через него — и к общей структуре объекта. На рис. 17, г этому случаю соответствует модель, формирующаяся при восприятии части 1 объекта сильно сфокусированным зрением. Весь процесс рассматривания объекта запечатлевается в серии кадров. В целом — это «фраза» изучения предмета, запечатленная в кадрах рецепторного поля.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава 9 ГИПОТЕЗА
Глава 9 ГИПОТЕЗА Формальная логика изучает не только формы абстрактного мышления, но и формы развития научного знания. Таковыми являются факты науки, рождающиеся из потребности объяснения фактов жизни, научная проблема, первоначальное решение проблемы в форме гипотезы,
Гипотеза
Гипотеза Гипотеза – это форма развития знаний, представляющая собой обоснованное предположение, выдвигаемое с целью выяснения свойств и причин исследуемых явлений.Можно выделить следующие черты, которые определяют гипотезу:1. Гипотеза – это необходимая форма для
Гипотеза
Гипотеза На пути от обнаружения научной проблемы к построению теории необходимым узловым, связующим пунктом выступает гипотеза — научное допущение или предположение, истинность которого еще требуется доказать.Необходимость гипотезы обусловливается тем, что законы
8. Гипотеза Большого Взрыва
8. Гипотеза Большого Взрыва Точно ответить на вопрос о происхождении Вселенной современная наука пока не может (и вряд ли будет в состоянии это сделать в ближайшее время – настолько он сложен). Однако у нее есть на этот счет более или менее обоснованные предположения.
5.14. Что такое гипотеза?
5.14. Что такое гипотеза? Слово гипотеза – греческое (hypothesis). Оно переводится на русский как «предположение». Гипотеза представляет собой предположение, как правило, научного характера, которое выдвигается с целью объяснения каких-либо объектов, явлений, событий и т. п. От
3. Ещё раз о механизмах общественного развития
3. Ещё раз о механизмах общественного развития В первой части работы я постарался показать, как однажды биосоциальным законам, которые управляли развитием популяций гоминидов, подобно тому, как это происходило во всех популяциях животных, ведущих общественный образ
§ 2. ГИПОТЕЗА И СЛЕДСТВЕННАЯ ВЕРСИЯ
§ 2. ГИПОТЕЗА И СЛЕДСТВЕННАЯ ВЕРСИЯ Почти всегда, когда человек начинает какое-либо исследование, он выдвигает предположение о его результатах, т.е. как бы видит желаемый результат в начале исследования. Такое предварительное решение вопроса, в большинстве случаев, служит
3. Гипотеза
3. Гипотеза Еще одной из важных форм развития знания выступает гипотеза (от греч. hypothesis — основание, предположение).Она неразрывно связана с предшествующими формами. Ее выдвижение так или иначе предполагает наличие фактов; а сама она направлена на решение той или иной
ДОГАДКА И ГИПОТЕЗА
ДОГАДКА И ГИПОТЕЗА Подобно тому как бабочка появляется на свет, только пройдя стадию гусеницы, так и теория рождается первоначально в виде гипотезы. Все наше теоретическое знание гипотетично по своему происхождению, а некоторая его часть — и по своему нынешнему
48. Гипотеза
48. Гипотеза Гипотезой называют предположение о каком-либо предмете или явлении, его причинах, связях, закономерностях природы, общества и государства, основанных на научных данных.Основанные на научных знаниях доказанные гипотезы можно называть научно обоснованными.
3. Гипотеза
3. Гипотеза Гипотезой называют предположение о каком-либо предмете или явлении, его причинах, связях, закономерностях природы, общества и государства, основанных на научных данных.Основанные на научных знаниях доказанные гипотезы можно называть научно обоснованными. Не
Гипотеза (Hypothése)
Гипотеза (Hypoth?se) Предположение, обычно выдвигаемое с экспериментальной или доказательной целью; идея, временно принимаемая за истинную с целью сделать из нее выводы и, в предельном случае, подтвердить или опровергнуть ее истинность. В экспериментальных науках гипотеза,