Приспособительный характер структур восприятия
Приспособительный характер структур восприятия
Человеческая способность восприятия является таким же результатом естественного отбора, как и любой другой признак организма. При этом селекция благоприятствовала в общем лучшему познанию объетивных черт того окружающего мира, в котором жили наши дочеловеческие предки.
(Shimony,1971,571)
Если наш познаваетельный аппарат был развит в эволюциооном приспособлении к окружающему миру, тогда этот факт должен обнаруживаться в определённых приспособлениях. Это обстоятельство особенно отчётливо видно на примере оптического восприятия. Распределение интенсивности солнечного света по различным длинам волн подчиняется планковскому закону (рис. 8, кривая 1). При температуре солнечной поверхности 5800 К, максимум распределения находится на уровне 510 нм.
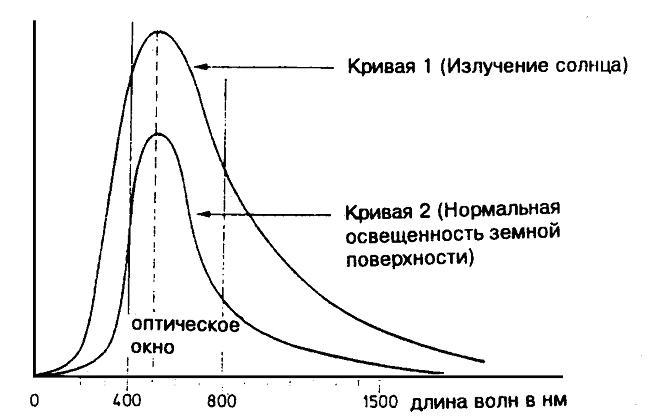
Рис. 8. Максимум интенсивности солнечного излучения находится в области видимого света (в зелёном).
Для этого излучения атмосфера проницаема только относительно. Так, рентгеновское и ультрафиолетовое излучение абсорбируется уже в высших, а инфракрасное в более близких к земле слоях атмосферы. Только для излучения между 400 и 800 нм (и для радиоволн) имеет атмосфера «окно» (рис. 8, кривая 2). Это окно практически совпадает с "оптическим окном" нашего восприятия (380 — 760 нм, см. стр. 46). Наш глаз восприимчив именно в том диапазоне, в котором электромагнитный спектр имет максимум. Согласно цветовому кругу на стр. 47 этот максимум находится в светлозелёном, примерно в центре цветового спектра.
Дело обстоит не так, что "как раз" видимый диапазон солнечного спектра проникает через нашу атмосферу. Наоборот, именно потому, что сравнительно небольшая часть широкого спектра солнечного излучения случайно оказалась в состоянии проникать через земную атмосферу, стала она для нас видимой областью этого спектра, «светом».
(v.Ditfurth, 1972,100)
Счастливой случайностью яявляется то, что оптическая прозрачность и механическая проницаемость практически совпадают (Cambell,1974,414). Твёрдые тела не прозрачны и не проницаемы, воздух и вода, напротив, прозрачны и проницаемы. Это согласование не действует при других длинах волн и, следовательно, представляет собой дополнительный селекционный фактор. Стекло и туман обладают в этом плане парадоксальными свойствами: стекло твёрдоё, но прозрачное, туман — наоборот. Однако стекло для эволюции не играло никакой роли, а туман только подчинённую роль.
Глаз, во всяком случае, настроен на оптимальное использование дневного света. До человеческой культуры солнце было ведь единственным действенным в селекции источником света; огонь, свет луны и звёзд имели малое значение. Также и у животных "оптическое окно" находится в этой же области. Оно может быть немного сдвинуто, как у пчёл (стр.48); но всегда используется благоприятная волновая область дневного света.
Эту констатацию не изменяет тот факт, что питон и гремучая змея, наряду с обычными глазами, имеют ещё и «инфракрасные», с помощью которых они «чувствуют» тепловое излучение идущее от теплокровных жертв; ибо эти глаза служат ведь не для видения дневного света(76).
Только это приспособление остаётся от старых представлений о том, что глаз, якобы, солнце-подобен, как у Плотина, метафизиков света или Гёте: был бы глаз не солнцеподобен, он не мог бы никогда взглянуть на солнце (Goethe: Zahme Xenien 3, und Einleitung zur Farbenlehre).
Не потому, что глаз изначально солнцеподобен, может он взглянуть на солнце, но потому что он сформировался в ходе миллиардов лет развития в мире, в котором реальное солнце уже вечно распространяет свои лучи перед глазами.
(Lorenz, 1943, 236)
Известно, что многие насекомые (пчёлы, стрекозы), некоторые рыбы, рептилии, птицы, обезъяны и люди могут различать цвета. По поводу эволюции цветового круга Шрёдингер уже в 1924 г высказал важное предположение(77):
Прежде всего воспринимался свет вообще, без всякой цветовой дифференциации.
Новая ступень развития была достигнута тогда, когда зрительный орган начал качественно различно реагировать на различные длины волн. Эта ступень двуцветности, по Шрёдингеру, соответствует жёлто-голубой дифференциации ощущений, как она проявляется также при частичной цветовой слепоте и у животных (насекомых). Переходным пунктом этой первой полярной дифференциации было ощущение белого цвета, который остаётся примитивнейшим признаком восприятия низшей ступени. К трёхцветности должет был вести дальнейший шаг, аналогичный полярному расколу белого на жёлтый и голубой. На этой третьей ступени жёлтый полярно распадается на ощущения красного и зелёного, точно так же, как на второй ступени белый распадается на жёлтый и голубой… Корни соразмерной восприятию простоты белого и жёлтого, а также наличие цветовой полярности, по Шрёдингеру, находятся в филогенезе зрительного органа. "Белый и жёлтый являются подлинными основными ощущениями, одно из одноцветной, другое из двуцветной стадии".
Это также объясняет, почему распространённым нарушением нормального цветовосприятия является красно-зелёная слепота, она соответствует атавизму дневного аппарата "первого рода". Полная цветовая идиферентность — атавизм "второго рода" — , правда, имеет место, но встречается реже, чем частая красно-зелёная слепота. Наиболее редким является выпадение восприятия голубого.
(Honl, 1954, 523f.)
То обстоятельство, что смешение всех цветов радуги кажется нам «белым» светом, точнее бесцветным, свидетельствует о приспособительном характере нашего цветовосприятия. Для аппарата восприятия было особенно биологически осмысленно интерпретировать нормальное освещение земной поверхности как нейтральное в цветовом отношении и только отклонения от нормальной освещённости осознавать как цвет.
Эволюционистские объяснения имеются также для спектральнорй восприимчивости светочувствительных клеток (волны различной днины, несмотря на одинаковую интенсивность раздражения, воспринимаются различно) и для цветоразрешающих способностей глаза.
Нижний порог восприимчивости фоторецептора в ретине находится в пределах единственного светового кванта. Но нервная система только тогда сообщает о световом ощущении, когда в течение короткого промежутка времени раздражаются многие соседние клетки. Это есть защитное средство против неизбежных помех и статистических колебаний, которые всегда проявляются в высокочувствительных приборах вследствие квантомеханической природы света. Если бы регистрировался каждый квант, то мы бы имели непрерывные бессистемные световые впечатления при отсутствии информационного содержания. Эти не имеющие значения сигналы отсекаются цензурой нервной системы.
Аналогично «фильтр» у ушей препятствует интерпретации как шорохов ударов молекул по барабанной перепонке, вследствие броуновского движения. Это очень напоминает утверждение Цицерона в "Somnium Scipionis" (De re publica) о том, что мы не можем воспринимать гармонию сфер, так как наши уши к ней слишком привыкли!
Прекрасным примером является временная разрешающая способность нашего сознания. Временной промежуток, который должен иметься между двумя событиями, чтобы они не воспринимались как одновременные, называют субъективным квантом времени (SZQ). У людей SZQ — около 1/16 секунды. Если друг за другом следует более 16 световых сигналов в секунду, наш глаз оказывается неспособным воспринимать их раздельно, они образуют впечатление светового потока. Этот факт используют в кино и телевидении, чтобы создавать непрерывные сцены и движение. Периодические звуковые раздражения, которые следуют со скоростью более 16 в секунду, воспринимаются как непрерывный звук. Аналогичное действует для касаний.
Информационная психология трактует SZQ как интервал, в который информационная единица (один бит) входит в краткосрочную память (Frank, 1970, 245).
У различных видов животных SZQ различно. Например, рыба-драчун нападает на собственное зеркальное отражение, если оно, с помощью особого устройства, демонстрируется более 30 раз в секунду; ниже этой частоты изображение не воспринимается как противник, у неё «рябит». Она обрабатывает, следовательно, большее число оптических сигналов в секунду. Таких животных образно называют "лупами времени". SCQ пчёл ещё существенно короче. Имели бы пчёлы кино, проектор у них должен был бы работать очень быстро.
Пчёлам нужно предъявлять более 200 изображений в секунду, чтобы у них не «рябило». Глаз пчёл может перерабатывать в одно время в 10 раз больше отдельных впечатлений, чем наш. Поэтому он блестяще приспособлен для восприятия движения, обработки быстро меняющихся впечатлений, хотя покоящиеся вещи в полёте проходят мимо.
(v. Frisch, 1969,83)
С другой стороны, SZQ улитки длиннее, чем 1/4 секунды. Палка, которая приближается к ней c такой скоростью, кажется ей покоящейся и она пытается её устранить. Она является, следовательно, "пожирателем времени".
Сенсорные системы животных приспособлены таким образом, чтобы получать такую информацию, которая важна для образа жизни их владельцев.
(Gregory,1972,229)
Этот приспособительный характер чувственного восприятия становится особенно отчёливым в ошибках и заблуждениях, которые проявляются в чуждом окружении. Лягушка умирает с голоду посреди дохлых мух, так как они не движутся (по поводу оптического восприятия лягушек см. стр. 44). В воде мы видим всё искажённо, так как наш глаз приспособлен к закону преломления в воздухе. Чтобы восстановить «нормальный» переход глаза-воздух мы должны использовать особые очки.
Аналогичным образом, барабанная перепонка настроена на большие амплитуды колебаний воздуха. В воде, где звуковые колебания имеют намного меньшую амплитуду, слышим мы поэтому много тише. Отсюда возникло ложное предположение, что рыбы якобы немые. В действительности, едва ли найдётся рыба, которая не издаёт звуков. Ввиду того, что дыхательный воздух водолаза под водой сильно концентрирован, его голос звучит гнусаво и сдавленно, как сообщают исследователи моря (напр., Cousteau). Но не только к плотности, но также к составу воздуха оказались приспособленными уши и голос.
Как известно, голос человека, говорящего в кислородно-гелиумной атмосфере, совершенно непроизвольно приобретает дребезжащее «мики-маусовское» звучание. В такой атмосфере, в которой гелий заменяет азот, прежде всего изменяется скорость звука. Тем самым изменяются резонансные свойства воздуха, которые при говорении в гортани преобразуются в колебания. Но структура нашей гортани приспособлена именно к свойствам нормальной атмосферы (v. Ditfurth, 1972, 373)
Приспособительный характер нашего трёхмерного пространственного восприятия, проявляется прежде всего в открытиях этологии, в том, что некоторые животные обладают худшим пространственным восприятием, чем мы.
Организмы из мало структурированных жизненных пространств нуждаются в менее точном и дифференцированном ориентировочном поведении, нежли те, которые на каждом шагу должны сталкиваться со сложными пространственными данностями. Наиболее гомогенным из всех жизненных пространств является океан, в котором имеются также отдельные свободно передвигающиеся существа, которые полностью лишены собственных ориентирующих реакций (напр., медузы.)
Если океан трёхмерен, то степь в определёенной степени двумерна. Среди степных птиц и млекопитающих имеются такие, которые не понимают вертикальных препятствий и не научаются их преодолевать.
(Lorenz,1954, 225f.)
Жители деревьев являются теми животными, которые в повседневной жизни овладевают сложнейшими пространственными структурами, и именно те из них, которые используют не когти, а хватающёю лапу. У них уже до прыжка в центральной нервной системе совершенно точно должно быть представлено не только направление, но также удаление, положение и форма цели. Ибо хватающая лапа должна сработать в правильном пространственном направлении и в точно определённый момент времени(78). Люди обладают относительно хорошим пространственным восприятием благодаря своим предкам, которые, живя на деревьях и используя хватающую лапу, приобрели хорошую центральную репрезентацию своего трёхмерно-структурированного окружения. Но этот факт ведёт непосредственно к дальнейшему предположению, которое в следующей главе формулируется как гипотеза.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Ракурсы восприятия
Ракурсы восприятия Кому из нас не случалось ощущать своё существование в самых разных масштабах? Я могу чувствовать себя отшельником в своей внутренней вселенной или жителем многолюдного города, патриотом своей страны или гражданином мира, участником исторических
11.2. Ленинские декреты и утверждение структур власти Советского государства
11.2. Ленинские декреты и утверждение структур власти Советского государства 11.2.1. Декрет о миреСразу же после первых слов о родившемся советском государстве с докладом о мире выступил Владимир Ильич Ленин. Он долго не мог начать речь из-за непрекращающегося скандирования
Структуры восприятия
Структуры восприятия a. Восприятие цветаКак уже вытекает из примеров на стр. 43, восприятие цвета является репрезентативным примером селективной и конструктивной функции нашего аппарата восприятия. Прежде всего, расположение цветов в (физическом) спектре и
Пригодность структур познания
Пригодность структур познания Независимо от собранных, наконец, результатов физиологических и психологических исследований, теория познания всегда пыталась определить соотношение опытной действительности и субъективного познания. При этом она, естественно, не
3.7. Сознание как один из уровней информационных структур
3.7. Сознание как один из уровней информационных структур Рассмотрим более подробно основополагающее философское понятие «сознание». Понятие «сознание» ввел в философию Р. Декарт (1596—1650 г.г.). По общепринятому в марксистской философии определению, сознание — это
§ 1. Историчность структур общественного сознания
§ 1. Историчность структур общественного сознания Возможен вопрос: кому принадлежит сознание? Если оно формируется в ходе человеческого взаимодействия, т.е. является почти буквально во взаимопонимании нескольких, двух как минимум, субъектов, так, может быть, оно вообще
2. Антиципации восприятия
2. Антиципации восприятия Принцип их таков: реальное, составляющее предмет ощущения, имеет во всех явлениях интенсивную величину, т. е. степень.ДоказательствоВосприятие есть эмпирическое сознание, т. е. такое сознание, в котором есть также ощущение. Явления как предметы
Обобщенность восприятия
Обобщенность восприятия Основная проблема моделирования сложных объектов для целей интеллекта — это преодоление избыточного разнообразия, борьба с избыточной информацией. Чем более развит интеллект, тем более подробными должны быть модели управляемых объектов, тем
«Круги» восприятия
«Круги» восприятия В предыдущем описании ФА был предложен наиболее легкий процесс распознавания моделей — по полному совпадению с эталонами. В действительности это не так. Сложные структуры, воспринятые рецепторами и запечатленные в кратковременной памяти как
Глава четвертая. К проблематике ноэтически-ноэматических структур
Глава четвертая. К проблематике ноэтически-ноэматических структур § 97. Гилетические и ноэтические моменты в качестве реальных, ноэматические — в качестве нереальных моментов переживания Вводя различение ноэтического и ноэматического, мы в предыдущей главе
"ФЕНОМЕНОЛОГИЯ ВОСПРИЯТИЯ"
"ФЕНОМЕНОЛОГИЯ ВОСПРИЯТИЯ" "ФЕНОМЕНОЛОГИЯ ВОСПРИЯТИЯ" ("Phenomenologie de la perception". Paris, 1945) - основное произведение Мерло-Понти, в котором исследуются проблемы специфичности существования экзистенции (см. Экзистенция) и ее отношений с миром как "жизненной коммуникации", как
Механизмы восприятия
Механизмы восприятия Языковое высказывание фиксируется в своей однозначности ситуацией, в которой оно сформулировано. Представим себе полутемную пещеру, а в ней двух человек. Один из них начинает говорить, первое слово звучит: «Panie…» («Пан…»). Второй может подумать, что
Фрагментарность восприятия
Фрагментарность восприятия Валентин Фёдорович Турчин в своей книге «Феномен науки: Кибернетический подход к эволюции» одну из глав назвал «Мир глазами лягушки». В ней он рассматривает один из самых удивительных феноменов восприятия.Глаз лягушки устроен и работает
Роль и средства орденских структур
Роль и средства орденских структур Опыт деятельности и алгоритмика «ликвидации» ордена тамплиеров, дошедшие до нас благодаря небывалой «засветке» этого ордена в период надежд «мировой закулисы» на католическое европейское единство — особо интересен для понимания
Иконография иконных структур повествования
Иконография иконных структур повествования Несомненной сугубой программностью в рамках общей программности сборника обладает статья собственно редактора и автора концепции сборника А.Л. Баталова «К интерпретации архитектуры собора Покрова на Рву (о границах