7.4. Цитоскелет и микротрубочки
7.4. Цитоскелет и микротрубочки
Если мы вдруг вообразим, что сложное поведение животных управляется только лишь нейронами, то скромная парамеция поставит нас перед фундаментальной проблемой. Эта инфузория перемещается по своему пруду с помощью многочисленных крохотных волосообразных конечностей — ресничек, — преследуя бактерий, которыми она питается и которых обнаруживает посредством различных внутренних механизмов, или отступая от возможной опасности, готовая мгновенно устремиться прочь. Она также может преодолевать препятствия, огибая их. Более того, парамеция, по всей видимости, способна обучаться на собственном опыте{88} — хотя эта наиболее замечательная ее способность некоторыми учеными оспаривается{89}. Как же все это может проделывать существо, не имеющее ни единого нейрона и синапса? В самом деле, поскольку вся парамеция — это всего лишь одна, пусть и большая, клетка, и притом не нейрон, ей просто негде все перечисленные способности разместить (см. рис. 7.2).
Несомненно, поведение парамеции — да собственно и прочих одноклеточных организмов, например, амеб — регулируется какой-то сложной системой управления, просто эта система построена не из нервных клеток. Ответственная за поведение парамеции структура, очевидно, является частью ее так называемого цитоскелета. Как можно предположить из названия, цитоскелет служит для поддержания формы клетки, однако у него имеются и многочисленные иные функции. Упоминавшиеся выше реснички представляют собой окончания волокон цитоскелета, но помимо них цитоскелет, похоже, содержит еще и собственно систему управления движением клетки, а также систему «конвейеров», осуществляющих транспортировку молекул внутри клетки. Словом, в единичной клетке цитоскелет выступает в роли этакой комбинации скелета, мускулатуры, конечностей, системы кровообращения и нервной системы.
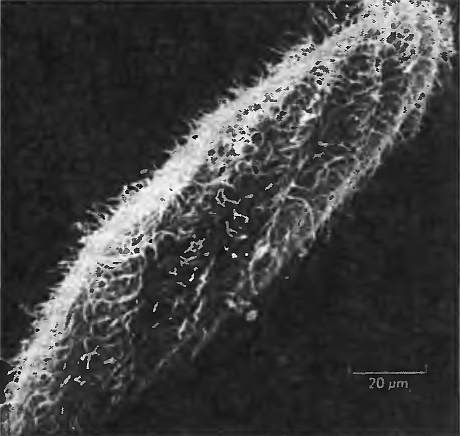
Рис. 7.2. Парамеция. Обратите внимание на волосообразные реснички, используемые для перемещения в воде. Они представляют собой наружные окончания цитоскелета парамеции.
Нас с вами в настоящий момент больше всего интересует, каким образом цитоскелет выполняет функции клеточной «нервной системы». Нейроны в нашем мозге сами являются отдельными клетками, причем у каждого нейрона есть свой собственный цитоскелет! Означает ли это, что в некотором смысле каждый отдельный нейрон располагает чем-то вроде «личной нервной системы»? Предположение весьма интригующее, и многие ученые склоняются к мнению, что нечто подобное действительно может иметь место. (См. первопроходческий труд Стюарта Хамероффа «Первичное вычисление: биомолекулярное сознание и нанотехнология» [183]; также рекомендую обратить внимание на статью [184] и многочисленные статьи в новом журнале «Нанобиология»[52].)
Прежде чем переходить к этим вопросам, необходимо рассмотреть вкратце общее устройство цитоскелета. Он состоит из протеиноподобных молекул, организованных в различного типа структуры: актин, микротрубочки и промежуточные волокна. Нас сейчас интересуют, главным образом, микротрубочки. Они представляют собой полые цилиндрические трубки с внешним диаметром около 25 нм и внутренним — около 14 нм (где «нм» обозначает «нанометр», т.е. 10—9 м), иногда организованные в более крупные трубкообразные волокна, состоящие из девяти дублетов, триплетов или частичных триплетов микротрубочек; в поперечном сечении такое волокно напоминает лопасти вентилятора, как показано на рис. 7.3, причем иногда по его центру также проходит пара микротрубочек. Как раз такое строение имеют реснички парамеции. Каждая микротрубочка представляет собой белковый полимер, состоящий из субъединиц, называемых тубулинами. Каждая субъединица тубулина, в свою очередь, представляет собой «димер», т.е. состоит из двух соединенных тонкой перемычкой частей, называемых ?-тубулин и ?-тубулин (приблизительно по 450 аминокислот в каждой). Эти, пары глобулярных белков, напоминающие по форме орех арахиса, уложены в слегка скошенную гексагональную решетку вдоль всей трубки, как показано на рис. 7.4. Обычно на каждую миктротрубочку приходится по 13 рядов димеров тубулина. Размеры димера составляют приблизительно 8 нм ? 4 нм ? 4 нм, а его атомное число — около 11 ? 104 (т.е. в одном димере содержится такое количество нуклонов, что его масса в абсолютных единицах равна приблизительно 10—14).
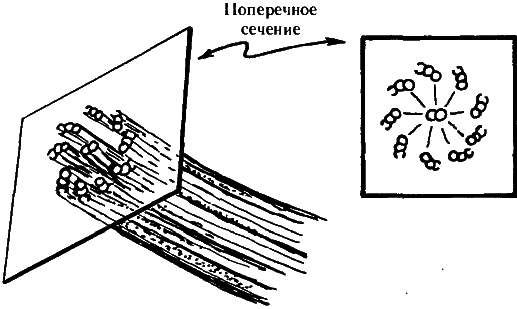
Рис. 7.3. Важной частью цитоскелета являются пучки крохотных трубочек (микротрубочек), организованных в структуры, напоминающие в поперечном сечении лопасти вентилятора. Такое строение имеют, например, реснички парамеции.
Димер тубулина может существовать в двух (по крайней мере) различных геометрических конфигурациях, называемых конформациями. В одной из таких конформаций молекулы тубулина располагаются под углом около 30° к оси микротрубочки. Есть основания полагать, что эти две конформаций соответствуют двум различным состояниям электрической поляризации димера, возникающим вследствие того, что электрон в центре перемычки ?-тубулин/?-тубулин занимает в различных конформациях различные положения.
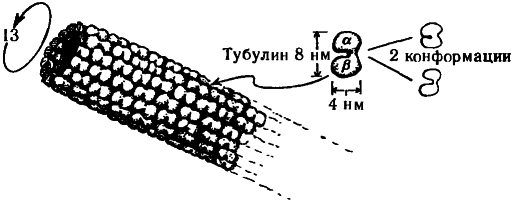
Рис. 7.4. Микротрубочка. Полая трубка, обычно состоящая из 13 рядов димеров тубулина. Каждая из молекул тубулина может существовать в двух (по крайней мере) конформациях.
«Центром управления» в цитоскелете является, по всей видимости, структура, называемая центром организации микротрубочек, или центросомой. Внутри центросомы имеется особая структура, называемая центриолью, которая состоит из двух цилиндрических волокон, по девять триплетов микротрубочек в каждом, образующих в пространстве структуру, похожую на «разделенную» букву «T» (см. рис. 7.5). (Цилиндрические волокна в общем аналогичны по структуре ресничкам, показанным на рис. 7.3.) Согласно Альбрехту-Бюлеру [7, 9], центриоль действует как глаз (!) клетки — идея чрезвычайно захватывающая, хотя и далеко еще не общепринятая. Какой бы ни была роль центросомы в нормальной, «повседневной», жизни клетки, она выполняет по крайней мере одну фундаментально важную задачу. На некоем критическом этапе она разделяется на две части, каждая из которых, по всей видимости, утягивает за собой пучок микротрубочек — хотя, пожалуй, точнее будет сказать, что каждая часть становится своего рода фокусом, вокруг которого и собираются микротрубочки. Эти микротрубочковые волокна каким-то образом связывают центросому с отдельными цепочками ДНК в ядре (в центральных точках, называемых центромерами), и цепочки ДНК расходятся — начиная тем самым удивительный процесс, известный специалистам под названием митоз, что означает всего-навсего деление клетки (см. рис. 7.6).

Рис. 7.5. Центриоль (по некоторым предположениям, глаз клетки) состоит из двух пучков микротрубочек (очень похожих на те, что изображены на рис. 7.3), образующих «разделенную» букву «T».
Рис. 7.6. При митозе (делении клетки) хромосомы разделяются, растаскиваемые пучками микротрубочек.
Может показаться странным, что внутри одной клетки действуют две столь разные «штаб-квартиры». Одна из них — ядро, где хранится основной генетический материал клетки, определяющий ее наследственность и уникальность, а также управляющий производством белкового материала, из которого, собственно, «строится» клетка. Другой управляющий центр — центросома с центриолью в качестве основного компонента, являющаяся, по всей видимости, главным узлом цитоскелета — структуры, которая, опять же по всей видимости, контролирует движение клетки и ее пространственную организацию. Предполагается, что присутствие этих двух различных «центров» в эукариотических клетках (клетках всех животных и почти всех растений на нашей планете, за исключением бактерий, сине-зеленых водорослей и вирусов) является результатом древней «инфекции», распространившейся по миру несколько миллиардов лет назад. Клетки, населявшие Землю прежде, были прокариотическими; они существуют и поныне в виде бактерий и сине-зеленых водорослей, и у них нет цитоскелета. Согласно одному из предположений [332], часть древнейших прокариот оказались каким-то образом связаны (возможно, «инфицированы») с неким видом спирохет (бактерий, перемещающихся с помощью нитеобразного хвоста, состоящего из цитоскелетных белков). Эти чуждые друг другу организмы постепенно «научились» жить вместе в симбиотической связи как единые эукариотические клетки. Так «спирохеты» превратились, в конечном счете, в цитоскелеты клеток — со всеми вытекающими последствиями для будущей эволюции, среди которых мы с вами!
Организация микротрубочек млекопитающих представляет интерес с математической точки зрения. На первый взгляд, число 13 не имеет какого-либо особого математического значения, однако это не совсем так. Оно принадлежит к знаменитой последовательности чисел Фибоначчи:
0, 1, 1, 2, 3, 5, 8, 13, 21, 34, 55, 89, 144, …
где каждое последующее число получается сложением двух предыдущих. Это может показаться случайным совпадением, однако хорошо известно, что числа Фибоначчи в биологических системах не редкость (и в гораздо более крупном масштабе). Например, в еловых шишках, цветках подсолнечника и пальмовых стволах наблюдаются спиральные или винтовые структуры с взаимопроникновением левых и правых закручиваний, причем количество рядов, закрученных в одном направлении, и количество рядов, закрученных в другом направлении, суть два соседних числа Фибоначчи (см. рис. 7.7). (Если внимательно рассмотреть такую структуру от одного конца до другого, можно обнаружить «место перехода», где числа рядов сменяются на следующую пару соседних чисел Фибоначчи.) Любопытно, что гексагональный узор микротрубочек демонстрирует очень похожую особенность — в общем случае даже еще более точно, — причем состоит этот узор (по крайней мере, обычно) из 5 правых и 8 левых винтовых структур, как показано на рис. 7.8. На рис. 7.9 я попытался изобразить, как такие структуры могли бы «выглядеть» изнутри микротрубочки. Число 13 выступает здесь как общее количество витков в спирали: 5 + 8. Любопытно также, что в двойных микротрубочках, встречающихся достаточно часто, внешний слой составной трубки обычно содержит 21 ряд димеров тубулина — следующее число Фибоначчи! (Не стоит, впрочем, чересчур увлекаться подобными построениями; например, в пучках микротрубочек в ресничках и центриолях бывает и по 9 рядов димеров — число, определенно не принадлежащее последовательности Фибоначчи.)

Рис. 7.7. Цветок подсолнечника. Как и во многих других растениях, отчетливо наблюдаются числа Фибоначчи. Во внешней области круга имеем 89 спиралей, закрученных по часовой стрелке, и 55 спиралей, закрученных против часовой стрелки. Ближе к центру появляются другие числа Фибоначчи.

Рис. 7.8. Представим, что микротрубочка разрезана вдоль и затем развернута в полосу. Можно видеть, что молекулы тубулина располагаются вдоль наклонных линий, причем каждый новый виток смещен относительно предыдущего на 5 или 8 молекул (в зависимости оттого, куда наклонена линия, вправо или влево).
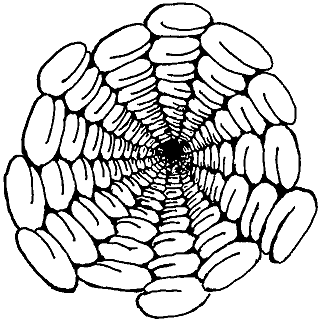
Рис. 7.9. Заглянем внутрь микротрубочки! Можно наблюдать спиральную структуру молекул тубулина 5 + 8.
Откуда в структуре микротрубочек берутся числа Фибоначчи? Относительно еловых шишек, цветков подсолнечника и т.д. существует несколько вполне убедительных теорий — кстати, среди тех. кто серьезно занимался этим вопросом, был Алан Тьюринг (см. [198], с. 437). Однако к случаю микротрубочек эти теории, вполне возможно, неприменимы, и для такого уровня следует искать какие-то другие объяснения. Коруга [228] высказал предположение, что числа Фибоначчи в структуре микротрубочки повышают эффективность ее как «информационного процессора». В самом деле, согласно Хамероффу с коллегами (которые пытаются нам это втолковать вот уже более десяти лет{90}), микротрубочки могут действовать как клеточные автоматы, передавая и обрабатывая сложные сигналы в виде волн различных состояний электрической поляризации молекул тубулина. Вспомним, что димеры тубулина могут существовать в двух (по крайней мере) различных конформационных состояниях и способных переходить из одного состояния в другое; последнее, очевидно, обусловливается сменой электрической поляризации молекулы на альтернативную. На состояние каждого димера воздействуют состояния поляризации каждого из шести его соседей (вследствие ван-дер-ваальсовых взаимодействий между ними), т.е. существуют вполне конкретные правила, определяющие конформацию каждого димера через конформации его соседей. Благодаря этому обстоятельству, каждая микротрубочка способна осуществлять передачу и обработку любого рода сообщений. С распространением сигналов, похоже, как-то связана транспортировка различных молекул вдоль микротрубочек, а также всевозможные соединения между соседними микротрубочками в виде своеобразных белковых «мостиков» — так называемые MAP (от microtubule associated proteins[53]); см. рис. 7.10. Коруга доказывает, что в случае структуры с числами Фибоначчи, подобной той, что реально наблюдается в микротрубочках, информация обрабатывается особенно эффективно. Должно быть, для такой организации микротрубочек и в самом имеется серьезная причина, поскольку, несмотря на некоторый разброс в числах, наблюдаемый в эукариотических клетках вообще, микротрубочки почти всех млекопитающих составлены именно из 13 рядов димеров.

Рис. 7.10. Микротрубочки обычно соединяются друг с другом посредством «мостиков» из так называемых белков, ассоциированных с микротрубочками (MAP).
Для чего микротрубочки нейронам? Каждый отдельный нейрон имеет свой цитоскелет. Какова его роль? Я уверен, что будущим исследователям предстоит сделать в этой области еще немало открытий, однако кое-что мы знаем уже сейчас. В частности, микротрубочки нейронов могут быть очень и очень длинными (по сравнению с диаметром нейрона, который составляет лишь 25—30 нм) — до нескольких миллиметров или даже длиннее. Более того, в зависимости от обстоятельств они способны расти или сокращаться, а также транспортировать молекулы нейромедиаторов. Внутри аксонов и дендритов также имеются микротрубочки. Хотя, как правило, на всю длину аксона каждая отдельная микротрубочка не тянется, они образуют сообщающиеся сети, охватывающие всю клетку, соединяясь между собой посредством упоминавшихся выше MAP-мостиков. Микротрубочки, по-видимому, ответственны за поддержание интенсивности синапсов и, несомненно, за изменение этой интенсивности в случае необходимости. Более того, они, похоже управляют ростом новых нервных окончаний, направляя их к точкам соединений с другими нервными клетками.
Поскольку после окончательного формирования мозга деление нейронов прекращается, необходимости в этой функции центросомы здесь нет. В центросомах нейронов, расположенных вблизи ядра, часто вовсе нет центриолей. Микротрубочки тянутся от центросом к окрестности пресинаптических окончаний аксона, а также в другую сторону, к дендритам и, через сокращающиеся актиновые нити, к дендритным шипикам, часто образующим постсинаптические окончания синаптической щели. Эти шипики способны расти и вырождаться, что, по-видимому, является существенным элементом общей пластичности мозга, благодаря которой система взаимных соединений в мозге подвергается непрерывным тонким изменениям. Насколько мне известно, существуют убедительные экспериментальные свидетельства важной роли микротрубочек в управлении пластичностью мозга.
Упомянем еще об одном любопытном факте. В пресинаптических окончаниях аксонов содержатся некие ассоциированные с миктротрубочками вещества, «работа» которых связана с высвобождением нейромедиаторов, а молекулы весьма примечательны с геометрической точки зрения. Эти вещества — клатрины — строятся из белковых тримеров (так называемых клатриновых трискелионов), этаких полипептидных трехлучевых звезд. Объединяясь в молекулу клатрина, трискелионы образуют геометрически правильные структуры, идентичные по общему строению многоатомным молекулам углерода, называемым «фуллеренами» (а также «бакиболами», или «мячами Баки»[54]) из-за их внешнего сходства со знаменитыми геодезическими куполами, которые проектировал и возводил американский архитектор Бакминстер Фуллер{91}. Клатрины, впрочем, гораздо больше фуллереновых молекул, поскольку одному атому углерода в фуллерене соответствует в клатрине целый трискелион, состоящий из нескольких аминокислот. Те клатрины, что заняты в высвобождении нейромедиаторов в синапсах, имеют форму усеченного икосаэдра — всем нам знакомого многогранника, по образу и подобию которого делают современные футбольные мячи (см. рис. 7.11 и 7.12).
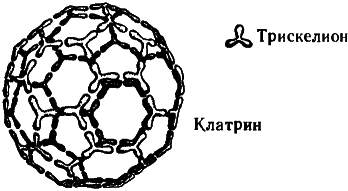
Рис. 7.11. Молекула клатрина (похожая общей структурой на фуллерен, но составленная не из атомов углерода, а из более сложных субструктур — белковых тримеров, называемых трискелионами). Изображенный на рисунке клатрин напоминает внешне обыкновенный футбольный мяч.
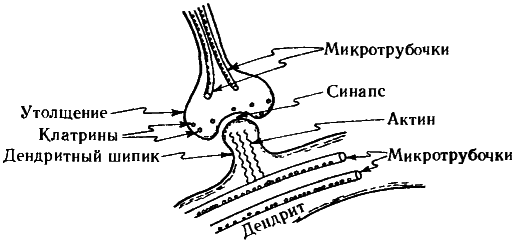
Рис. 7.12. Клатрины, подобные тому, что изображен на рис. 7.11, располагаются (вместе с окончаниями микротрубочек) в пресинаптическом утолщении аксона и, по всей видимости, участвуют в управлении интенсивностью синапса; также на интенсивность синапса влияют сокращающиеся актиновые нити в дендритных шипиках, управляемых микротрубочками.
В одном из предыдущих параграфов был поставлен важный вопрос: что управляет изменением интенсивности синапсов и определяет места размещения функционирующих синаптических связей? Учитывая имеющиеся свидетельства, можно уверенно предположить, что центральную роль в этих процессах играет цитоскелет. Как же это предположение может нам помочь в поиске невычислимой сущности разума? Пока что оно, похоже, говорит нам лишь о том, что потенциальная вычислительная мощность мозга оказывается гораздо большей, чем можно было бы ожидать, используй мозг в качестве простейших вычислительных блоков «цельные» нейроны.
В самом деле, если простейшими вычислительными блоками мы теперь будем считать димеры тубулина, то придется предположить, что потенциальная вычислительная мощность мозга просто неимоверно превосходит все то, что предполагали самые смелые теоретики от ИИ. Основываясь на «цельнонейронной» модели, Ханс Моравек в своей книге «Дети разума» [267] предположил, что человеческий мозг может в принципе достичь производительности порядка 1014 операций в секунду, но не более того; это при том, что в мозге имеется около 1011 функционирующих нейронов, каждый из которых способен посылать примерно по 103 сигналов в секунду (см. §1.2). Если же в качестве элементарного вычислительного блока взять димер тубулина, то следует учесть, что на каждый нейрон приходится около 107 димеров; соответственно, элементарные операции теперь выполняются где-то в 106 раз быстрее, в результате чего получаем 1027 операций в секунду. Возможно, производительность современных компьютеров и вправду уже начинает приближаться к первой цифре, 1014 операций в секунду (как весьма убежденно доказывают Моравек и его единомышленники), однако несмотря на все эти успехи, достичь в обозримом будущем производительности 1027 операций в секунду не представляется возможным.
Разумеется, можно смело утверждать, что мозг работает далеко не со стопроцентной «микротрубочковой» эффективностью, какую приведенные выше цифры предполагают. Тем не менее, ясно, что возможность «микротрубочкового вычисления» (см. [183]) позволяет совсем по-иному взглянуть на некоторые из аргументов в пользу неминуемого наступления эпохи искусственного интеллекта человеческого уровня. Можем ли мы теперь поверить хотя бы в то, что уже сегодня возможно{92} численно воспроизвести умственную деятельность червя нематоды, только потому, что мы вроде бы «закартографировали» и численно смоделировали его нервную систему? Как было отмечено в §1.15, умственные способности обычного муравья намного превосходят все то, что на настоящий момент реализовано посредством стандартных ИИ-процедур. Впору поинтересоваться, сколько же муравей выигрывает в производительности благодаря гигантскому массиву своих «микротрубочковых информационных нанопроцессоров», если сравнивать с тем, чего он смог бы добиться, располагай он лишь «переключателями цельнонейронного типа». Что до парамеции, то тут, как вы понимаете, оснований для предъявления иска нет.
Однако аргументы, представленные в первой части, предполагают гораздо более сильное заявление. Я утверждаю, что способность человека к пониманию выходит за рамки какой угодно вычислительной схемы. Если мозгом человека управляют микротрубочки, то в микротрубочковых процессах должно быть что-то принципиально отличное от простого вычисления. Я утверждал, что такая невычислимая активность должна быть следствием достаточно макроскопической квантовой когерентности, объединенной неким тонким образом с макроскопическим поведением — с тем, чтобы обеспечить возможность протекания в системе тех новых физических процессов, что придут на смену бытующей в современной физике паллиативной R-процедуре. В качестве первого шага мы должны выяснить, какова же подлинная роль квантовой когерентности в цитоскелетной активности.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
7.6. Микротрубочки и сознание
7.6. Микротрубочки и сознание Есть ли прямые свидетельства того, что феномен сознания в той или иной мере обусловлен деятельностью цитоскелета и, в частности, его микротрубочек? Как ни странно, есть. Причем получено оно путем обращения к проблеме сознания с неожиданной