Глава 10 БИОЛОГИЧЕСКИЕ ПАТТЕРНЫ ДВИЖЕНИЯ
Если попросить биолога определить понятие «жизнь», его ответ, вероятно, окажется таким: жизнь — это рост, обмен веществ, размножение, мутация. Помимо всего перечисленного, для большинства живых существ характерна еще и почти немыслимая координация между отдельными частями организма, что особенно ярко проявляется у животных и человека. Миллионы — если не миллиарды — различных клеток в высшей степени слаженно и организованно работают вместе, обеспечивая бесперебойную сердечную деятельность, кровообращение, дыхание и движение. То же относится и к осуществлению восприятия всех типов (в частности, человеческой мыслительной и речевой деятельности). Совершенно очевидно, что исследование такого рода взаимодействий естественным образом представляет собой одну из задач, стоящих перед синергетикой.
В самом деле, в последние десятилетия в этой области был достигнут немалый прогресс. При этом полностью оправдалась общая стратегия синергетики, нацеленная на изучение состояний, при которых происходят качественные изменения поведения исследуемой системы. Здесь представляется целесообразным начинать с постановки относительно простых вопросов. К таковым, на мой взгляд, может быть, например, отнесен вопрос о различных лошадиных аллюрах (шаг, рысь и галоп). При смене лошадью аллюра (представляющего собой, можно так сказать, двигательный стереотип, или паттерн движения) имеет место очевидный скачкообразный переход. Вопрос, с которого мы начнем, заключается в следующем: может ли такая смена аллюра расцениваться как фазовый переход?
Конечно, рассмотрение движения четырех конечностей — дело довольно-таки сложное; однако благодаря счастливому стечению обстоятельств, эта задача оказалась в значительной степени упрощена. В 1984 году мы встретились с американским физиологом Скоттом Келсо, и он сообщил мне о своем эксперименте, связанном с изучением паттернов движения. Поскольку этот эксперимент и его теоретическое обоснование стали отправной точкой для целого ряда исследований в этом направлении, я опишу его несколько подробнее. Речь идет об эксперименте, который легко сможет провести и сам читатель. Келсо предлагал испытуемым подвигать указательными пальцами обеих рук, следя за тем, чтобы пальцы двигались параллельно друг другу (рис. 10.1), причем ритм движения задавался метрономом.
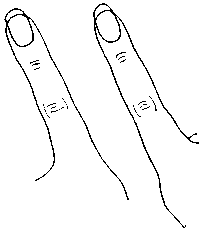
Рис. 10.1. Параллельное движение пальцев в эксперименте С. Келсо
Пока частота, задаваемая метрономом, была невелика, испытуемые хорошо справлялись с заданием, и параллельное положение пальцев при движении сохранялось. Однако как только частота ударов метронома повышалась, и испытуемые должны были увеличить и свою скорость движения, происходило следующее. При достижении определенной — индивидуальной для каждого испытуемого — скорости возникало качественное изменение, причем совершенно непроизвольное: параллельное положение пальцев сменялось симметричным (рис. 10.2 и 10.3). Совершенно очевидно, что здесь мы имеем дело с качественным макроскопическим изменением — совершенно в синергети-ческом духе, если можно так выразиться.

Рис. 10.2. Симметричное движение пальцев
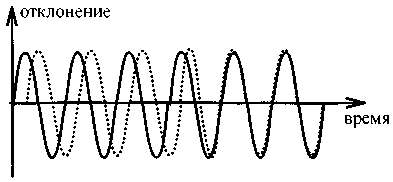
Рис. 10.3. Скорость метронома медленно повышается: отклонения правого (сплошная линия) и левого (пунктир) пальцев в зависимости от времени; совпадение кривых соответствует симметричному движению пальцев
Однако что же в этом примере играет роль контрольного параметра, и что здесь является параметром порядка? Да и возможно ли вообще, имея дело со столь сложной системой, как «человек», по-прежнему использовать эти понятия? Контрольный параметр, впрочем, отыскался быстро: им оказалось не что иное, как частота движения. На роль же параметра порядка, претерпевающего качественное изменение, напрашивается такая характеристика, как относительное положение пальцев. Чтобы перейти к корректному с математической точки зрения изложению, введем понятие так называемой относительной фазы, суть которого поясняется на рис. 10.4. В соответствии с этим понятием, параллельное движение пальцев соответствует фазе ? = 180°, а симметричное — фазе ? = 0°. Здесь следует упомянуть о двойном значении слова «фаза», так как из-за этого легко могут возникнуть недоразумения. С одной стороны, мы определяем положение пальцев относительно друг друга как фазу ?; с другой стороны, мы говорим о фазовом переходе при внезапном, скачкообразном изменении макроскопического состояния системы: например, при переходе от льда (твердое состояние) к воде (жидкое состояние) или при появлении цилиндрических структур в жидкости, нагретой до температуры, превышающей критическое значение. В дальнейшем словосочетание «фазовый переход» мы будем употреблять только в последнем значении; в остальных случаях мы будем говорить об «изменении фазы».
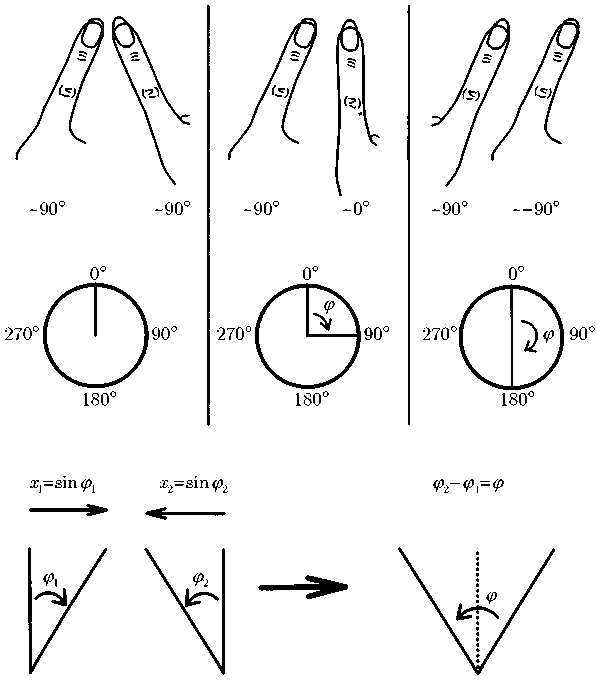
Рис. 10.4. На рисунке показано, как движение пальцев относительно друг друга можно передать через так называемую относительную фазу. Вверху слева направо: относительная фаза равна нулю; относительная фаза равна 90°; относительная фаза равна 180°. Внизу: зависимость между отклонениями пальцев x1, x2, фазами ?1, ?2 и относительной фазой
Как уже было показано в четвертой главе, фазовые переходы могут быть наглядно описаны при помощи так называемых холмистых ландшафтов (рис. 4.15-4.18). Такие ландшафты могут выглядеть и совершенно иначе, чем те, что представлены на наших иллюстрациях. Если рассматривать изменение фазы в экспериментах Келсо как фазовый переход, то нам следует сначала подобрать ландшафт, который деформировался бы в соответствии с изменениями фазы. В работе, опубликованной мною в сотрудничестве со Скоттом Келсо и Гербертом Бунцем, я предложил следующий ряд ландшафтов (рис. 10.5). В верхнем ряду слева изображена кривая, на которой имеются две точки устойчивого равновесия, расположенные в правой и левой верхних долинах; они соответствуют параллельному движению пальцев. При повышении скорости движения ландшафт изменяется: верхние долины, деформируясь, исчезают, и шарик скатывается в нижнюю, центральную долину; другими словами, значение фазы становится равно нулю, что соответствует уже не параллельному, а симметричному движению пальцев.

Рис. 10.5. Холмистый ландшафт для относительной фазы. При повышении скорости (частоты) движения пальцев — контрольного параметра — ландшафт деформируется, и верхние долины исчезают; при этом наблюдается обусловленная периодичностью движения взаимосвязь правой и левой верхних долин
Основываясь на данной модели, можно сделать целый ряд интересных выводов. Во-первых, перед нами пример так называемого гистерезиса.
Начнем рассмотрение с прежнего положения шарика в одной из верхних долин, точки устойчивого равновесия; пальцы испытуемых движутся параллельно. При увеличении скорости движения шарик в конце концов скатывается в центральную долину — в точку, в которой значение фазы равно нулю; пальцы движутся симметрично. Допустим, теперь скорость движения начнет снижаться, но пальцы по-прежнему будут двигаться симметрично; как бы ни снижалась скорость, спонтанного изменения положения пальцев не происходит, и причины становятся очевидны, стоит лишь взглянуть на нашу схему: шарик не может вернуться ни в одну из верхних долин, спонтанно «выпрыгнув» из центральной, самой глубокой, а потому — естественно — остается в ней. Другими словами, текущее положение шарика (положение пальцев относительно друг друга при одной и той же скорости движения) зависит от предшествующих его положений. При абсолютно одной и той же скорости шарик может находиться как в верхней долине (если мы, начав с небольшой скорости, постепенно увеличиваем ее), так и в нижней, центральной (в том случае, если скорость движения пальцев уменьшается). Этот феномен носит название гистерезис. Для испытуемых это означает два совершенно различных типа движения при одном и том же значении контрольного параметра «скорость движения пальцев».
Когда при деформации ландшафта исчезают верхние долины, мы можем наблюдать еще два важных феномена, предсказанных нами при рассмотрении подобных ландшафтов в пятой главе. Речь идет о критических флуктуациях и критическом замедлении. То есть, вблизи критической точки перехода от параллельного движения к симметричному можно ожидать сильных флуктуаций относительной фазы. Уточним: при больших количествах повторений эксперимента вдали от точки перехода постоянно наблюдается одна и та же фаза; вблизи же этой точки эксперимент каждый раз дает разные результаты. На рис. 10.6 даны результаты, полученные Келсо: график дает наглядное представление о том, насколько сильны флуктуации. Для наблюдения критического замедления, Келсо намеренно сбивал частоту движения пальцев и отмечал, сколько времени потребуется испытуемому для восстановления прежнего ритма движения; при этом скорость изменения частоты метронома, задающей ритм движения пальцев, варьировалась. Таким образом, и в экспериментах Келсо были обнаружены феномены, уже известные синергетике из исследований поведения разных других систем (например лазера); впоследствии результаты этих экспериментов были подтверждены рядом других ученых.

Рис. 10.6. По вертикали отложена амплитуда колебаний относительной фазы, по горизонтали — частота движения пальцев
После этого было проведено еще несколько экспериментов, направленных на изучение паттернов движения; например, исследовалась корреляция между движением локтя и запястья. В высшей степени интересными оказались эксперименты по исследованию корреляции между движениями двух сидящих рядом людей, которые должны были качать ногой симметрично друг другу: при увеличении скорости ноги испытуемых начинали вдруг двигаться параллельно. Количественные измерения показывают, что переход совершается в полном согласии с уравнениями нашей теории (рис. 10.7).
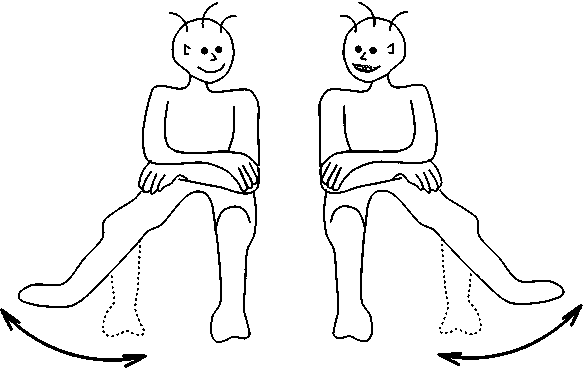
Рис. 10.7. Фазовый переход может произойти и при скоординированном движении ног двух испытуемых
Почему же, собственно, эти эксперименты представляют такой интерес для биологов? Да потому, что они имеют отношение к очень важному для биологии вопросу: каким образом осуществляется управление движением отдельных конечностей. Может быть, в мозге — как, например, в компьютере — имеется некая двигательная программа, которая в нужный момент просто переключается с одного типа управления на другой? Или все же перед нами — читатель об этом уже, наверное, догадывается — феномен самоорганизации? Все три обнаруженных эффекта — гистерезис, критические флуктуации и критическое замедление — совершенно не объяснимы при рассмотрении их в рамках «компьютерной» модели управления движением. Чего ради компьютер в нашей голове (пусть мы даже допустим его существование) будет изменять свою программу в зависимости от предыстории движения? Более того, компьютер, работающий на основе абсолютно детерминистских принципов, просто не в состоянии выдавать переменные результаты при одинаковых исходных данных! Результаты экспериментов значительно лучше согласуются с предположением о том, что здесь имеет место феномен самоорганизации — точно так же, как возникновение когерентного лазерного света является воплощением эффекта самоорганизации отдельных атомов лазера. В случае же биологического процесса речь может идти о самоорганизации нейронной сети, связанной с мышцами и клеточными тканями. Естественно, это уже совершенно новая точка зрения на происходящее, и именно ее следует выдвинуть на первый план.
Впрочем, и раньше существовали некоторые указания на то, насколько важны для движения конечностей эффекты самоорганизации. Несколько десятилетий назад, например, знаменитый физиолог Эрих фон Хольст провел следующий эксперимент. Он ампутировал многоножке все конечности за исключением шести, после чего животное это начало двигаться подобно насекомому; затем число конечностей было уменьшено до четырех, и многоножка передвигалась на них подобно любому четвероногому, например, лошади. Невозможно представить, что крошечный мозг многоножки способен содержать «двигательные программы» для всех возможных комбинаций конечностей, оставшихся после ампутаций; мы в состоянии объяснить этот феномен, только предположив возможность самоорганизации. Похожие результаты были получены русским физиологом М. Л. Шиком в ходе экспериментов с так называемыми децеребрированными кошками. Несмотря на то, что у этих кошек был удален мозжечок, они оставались в состоянии бежать по движущейся дорожке. При электрическом раздражении спинного мозга кошка двигалась шагом или переходила на галоп — «аллюр» зависел от силы раздражения. Совершенно очевидно, что постоянное раздражение выступает в данном случае в роли контрольного параметра, который побуждает нервную систему животного к самоорганизации, в результате чего возникает определенный паттерн движения. Похоже, синергетика открывает совершенно новые перспективы для исследования управления двигательными функциями.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК